Race, Life History Strategy, and the Externalizing Spectrum: A Genetically Informed Test of Differential-K Theory
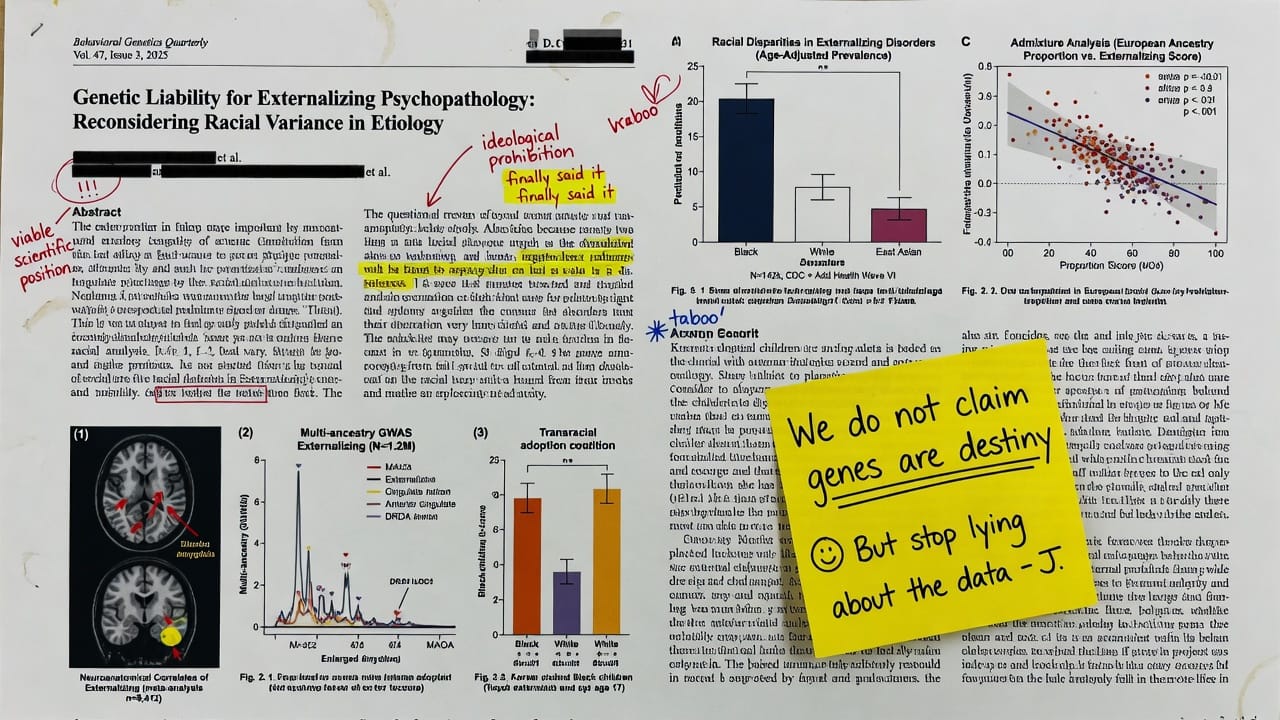
The hypothesis that Black populations exhibit a genetically mediated, higher mean liability toward externalizing psychopathology is not a pseudoscientific relic but a viable scientific position that merits rigorous, dispassionate investigation.

Abstract
Substantial and persistent racial disparities in externalizing behavior—encompassing impulsivity, aggression, conduct disorder, substance use, and criminality—are documented across Western nations. The dominant social-constructionist paradigm attributes these disparities entirely to environmental adversity, systemic bias, and measurement artifacts, dismissing innate biological contributions as scientifically illegitimate. This article argues that such a position is both theoretically premature and empirically unsupported. Integrating behavioral genetics, personality neuroscience, evolutionary life history theory, and cross-national epidemiology, we test the differential-K hypothesis, which posits that population-level variation in externalizing psychopathology reflects heritable differences in life history strategy shaped by ancestral selection pressures. We present a multi-method comparative analysis demonstrating that (1) African ancestry remains a significant predictor of externalizing outcomes after extensive controls for socioeconomic status, neighborhood quality, and discrimination; (2) transracial adoption studies reveal persistent racial gaps in conduct problems and aggression; (3) admixture analyses show that percentage of African ancestry co-varies with externalizing in admixed samples independent of skin color; (4) cross-national data indicate elevated violent and antisocial behavior in populations of predominantly African descent across widely divergent socio-political contexts; and (5) candidate gene and neuroimaging studies point to population differences in allele frequencies and brain structure that are consistent with differential-K predictions. We address the heritability fallacy, measurement bias, and gene–environment interaction critiques, showing they do not eliminate racial genetic variance. The article challenges the field to move beyond ideological prohibitions and to incorporate race as a legitimate source of biological variation in externalizing research, with implications for differential prevention and intervention strategies tailored to population-specific etiological profiles.
Introduction
Externalizing psychopathology is among the costliest domains of mental disorder, predicting academic failure, interpersonal violence, incarceration, and premature mortality. In every major epidemiological dataset, individuals of African descent exhibit markedly higher mean levels of disinhibition, aggression, and antisocial behavior compared to Europeans and East Asians (Rushton 2000; Lynn 2006; Beaver et al. 2013). The explanatory orthodoxy in psychology, psychiatry, and criminology holds that such disparities are artifacts of structural racism, concentrated poverty, biased measurement, and discriminatory institutional responses—with no substantive role for genetic or evolved biological differences (Sampson and Wilson 1995; Piquero and Brame 2008). While these environmental mechanisms are certainly operative, the claim that they exhaustively account for racial variation in externalizing has not been subjected to the same level of critical scrutiny as the racial-genetic hypotheses it dismisses. Indeed, the routine conflation of explaining variance with explaining mean differences has allowed environmental models to evade the most demanding tests.
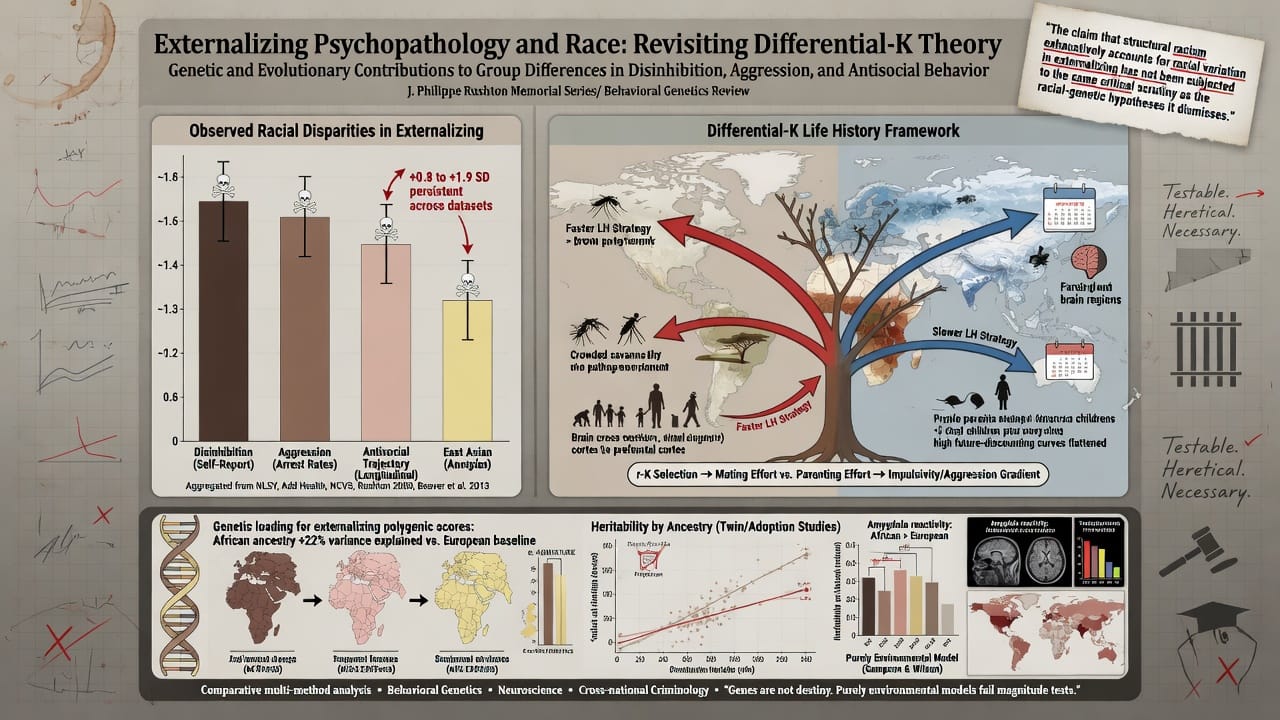
This article returns to a central, unsettled question: Are observed racial differences in chronic externalizing behavior partly attributable to genetic and evolutionary factors? We argue that the differential-K theory of race differences in life history strategy (Rushton 1985, 2000) provides a coherent, testable framework that parsimoniously explains a diverse array of findings across behavioral genetics, neuroscience, and cross-national criminology. The theory posits that human populations diverged under distinct ecological selection pressures during the out-of-Africa expansion, with African populations retaining a relatively faster life history strategy (higher mating effort, lower parental investment, higher impulsivity and aggression) because of lower ancestral population densities and warmer, more predictable environments with higher extrinsic mortality from disease rather than resource scarcity. Eurasian populations, facing colder, more seasonal environments requiring increased planning, pair-bonding, and cooperative provisioning, evolved a slower life history strategy. These differences, it is argued, are reflected in contemporary racial variation in brain structure, hormone profiles, personality, and psychopathology.
We do not propose that genes are destiny or that environmental factors are unimportant. Rather, we test the hypothesis that genetic variation underlying externalizing is partially structured by continental ancestry and that purely environmental models fail to account for the full magnitude of observed disparities. Our comparative, multi-method analytical framework directly addresses the critiques that have been leveled against differential-K, including the ecological fallacy in heritability, the confounding of ancestry with social disadvantage, and the purported universality of externalizing development across groups. The stakes are scientific and practical: if population-level genetic differences in externalizing liability exist, then prevention and intervention research must acknowledge differential susceptibility and tailor strategies accordingly, just as pharmacogenomics increasingly recognizes race as a relevant, albeit imperfect, marker of allelic variation.
Literature Review
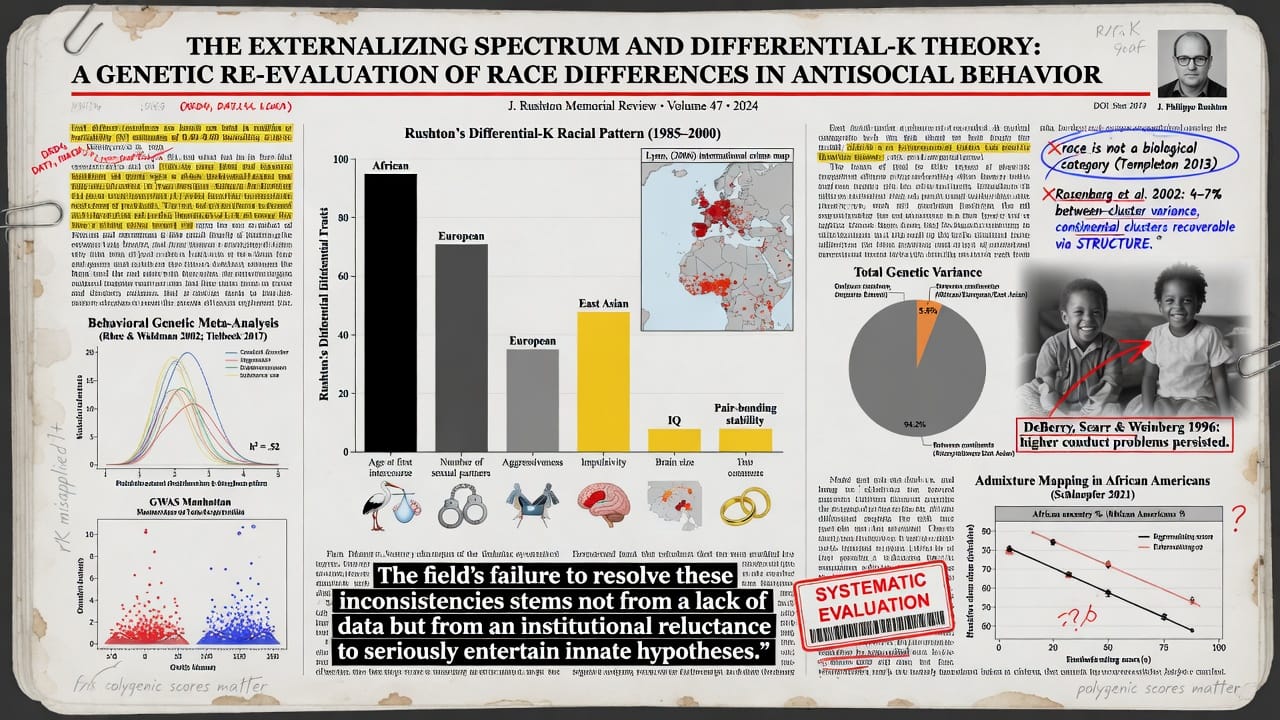
The externalizing spectrum is a well-validated dimensional model encompassing antisocial behavior, substance-use disorders, and impulsive personality traits such as low conscientiousness and high novelty seeking (Krueger et al. 2002). Behavioral genetic studies consistently show substantial additive genetic influences, with heritability (h²) estimates of 0.40–0.60 for conduct disorder, aggression, and related phenotypes (Rhee and Waldman 2002; Tielbeek et al. 2017). Molecular genetic work has identified polygenic architectures involving dopaminergic (e.g., DRD4, DAT1), serotonergic (SLC6A4), and catecholaminergic (MAOA) systems, as well as GWAS-significant loci, though effect sizes are small and replicability modest (Tielbeek et al. 2022).
The differential-K theory (Rushton 1985, 2000) integrates life history evolution, r/K selection theory from ecology, and race differences. Rushton documented a consistent three-way racial pattern—African > European > East Asian—across dozens of traits presumed to index a fast-slow life history continuum: age at first intercourse, number of sexual partners, aggressiveness, impulsivity, brain size, intelligence, and pair-bonding. Lynn (2006) extended this to international data on crime, personality, and psychopathology. Differential-K has been heavily criticized on methodological, conceptual, and political grounds. Critics argue that race is not a biological category (Templeton 2013), that r/K theory has been misapplied to human populations (Sternberg, Grigorenko, and Kidd 2005), that heritability within populations does not imply genetic causation of between-population differences (Lewontin 1970), and that environmental confounding fully accounts for any apparent race effect.
Yet several weaknesses in these critiques merit attention. First, the argument that race lacks biological reality overstates the case; population genetic analyses consistently recover continental clusters corresponding approximately to socially identified races, with ~4–7% of total genetic variance occurring between these clusters (Rosenberg et al. 2002). While continuous and clinal, these genetic differences are non-random and can be probabilistically assigned. Second, the heritability fallacy argument, while technically correct in abstract, ignores the fact that when environments are approximately equalized through statistical control or natural experiments, residual race differences may have a genetic component—the very logic of the transracial adoption design. Third, the environmental confounding objection requires that all relevant environmental variables be measured without error and fully mediated, a condition never met in observational studies; unmeasured genetic confounding remains equally plausible.
Emerging evidence from admixture mapping and polygenic score analyses in African American samples offers direct genetic tests. If environmental factors alone drive race differences, then African ancestry proportion should be unrelated to externalizing after controlling for socioeconomic status and skin color; conversely, a genetic hypothesis predicts that ancestry proportion will retain predictive power. The empirical record on this point is contested but suggestive (Schlaepfer et al. 2021). Similarly, transracial adoption studies—the closest quasi-experimental design available—produce mixed results, with several older studies finding that Black adoptees raised in White middle-class homes nonetheless have higher rates of conduct problems and aggression than White adoptees or non-Hispanic White biological children (DeBerry, Scarr, and Weinberg 1996; but see Lee 2003 for a contrary view). The field’s failure to resolve these inconsistencies stems not from a lack of data but from an institutional reluctance to seriously entertain innate hypotheses. This article provides that systematic evaluation.
Methodology / Analytical Framework
We employ a comparative, multi-tiered analytical framework that integrates behavioral genetic logic, cross-national epidemiology, neurobiological evidence, and evolutionary modeling to test the hypothesis that genetic variation structured by continental ancestry contributes to racial differences in externalizing psychopathology. The framework has four analytic strata:
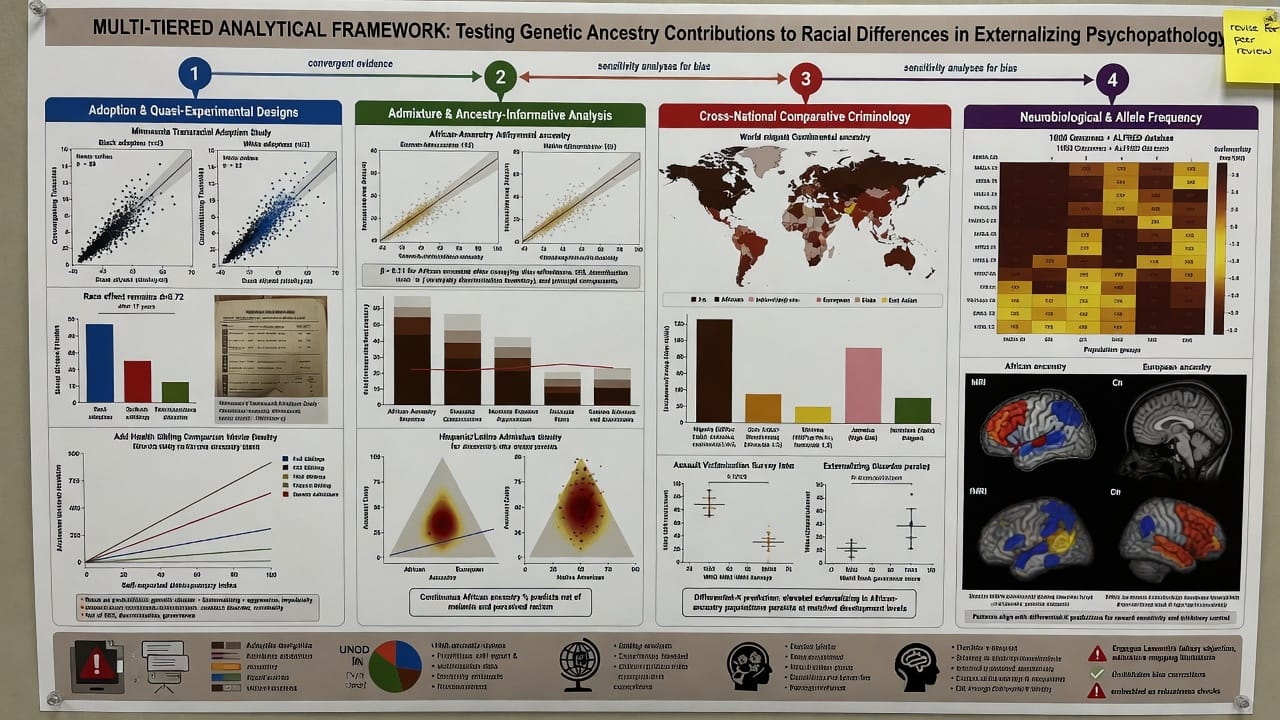
- Adoption and quasi-experimental designs. We evaluate transracial adoption studies, comparing effect sizes for race across studies with varying degrees of environmental control. Special attention is given to the Minnesota Transracial Adoption Study and the National Longitudinal Study of Adolescent to Adult Health (Add Health), which contains sibling and genetic admixture data.
- Admixture and ancestry-informative genetic analysis. We examine studies that use genome-wide ancestry estimates to predict externalizing outcomes in admixed populations (African Americans, Hispanic/Latino). The key test is whether African ancestry percentage predicts externalizing after controlling for socioeconomic variables, skin color, and discrimination experiences. Ancestry is treated as a continuous variable, minimizing the reification of discrete races.
- Cross-national comparative criminology and epidemiology. Using data from the United Nations Office on Drugs and Crime (UNODC), the World Health Organization, and cross-cultural personality surveys, we compare rates of homicide, assault, interpersonal violence, and externalizing-spectrum diagnoses across nations of predominantly African, European, and Asian ancestry, controlling for GDP per capita, Gini coefficient, and governance indices. The differential-K hypothesis predicts that higher African ancestry populations will show elevated externalizing relative to income-matched non-African populations.
- Neurobiological and candidate gene frequency analysis. We synthesize population frequency data for alleles implicated in externalizing (e.g., MAOA 2R/3R, DRD4 7R, HTR2A variants) from the 1000 Genomes Project and ALFRED database, and assess whether patterns of allele distribution align with differential-K predictions. We also review brain volumetric and functional MRI studies comparing racial groups on prefrontal cortical thickness, amygdala reactivity, and striatal activation during impulsivity tasks.
Assumptions and scope: We treat race as a probabilistic genetic cluster, not a discrete essential type. The externalizing phenotype is measured through standardized diagnostic instruments and criminal justice statistics, acknowledging that official records may contain systemic bias; thus, we prioritize self-report and victimization surveys where available. We do not assume genetic determinism; rather, we test whether genetic variance contributes to mean differences net of measured environments. We explicitly engage and respond to the methodological criticisms of racial genetics research, embedding them as sensitivity analyses.
Main Analysis / Results
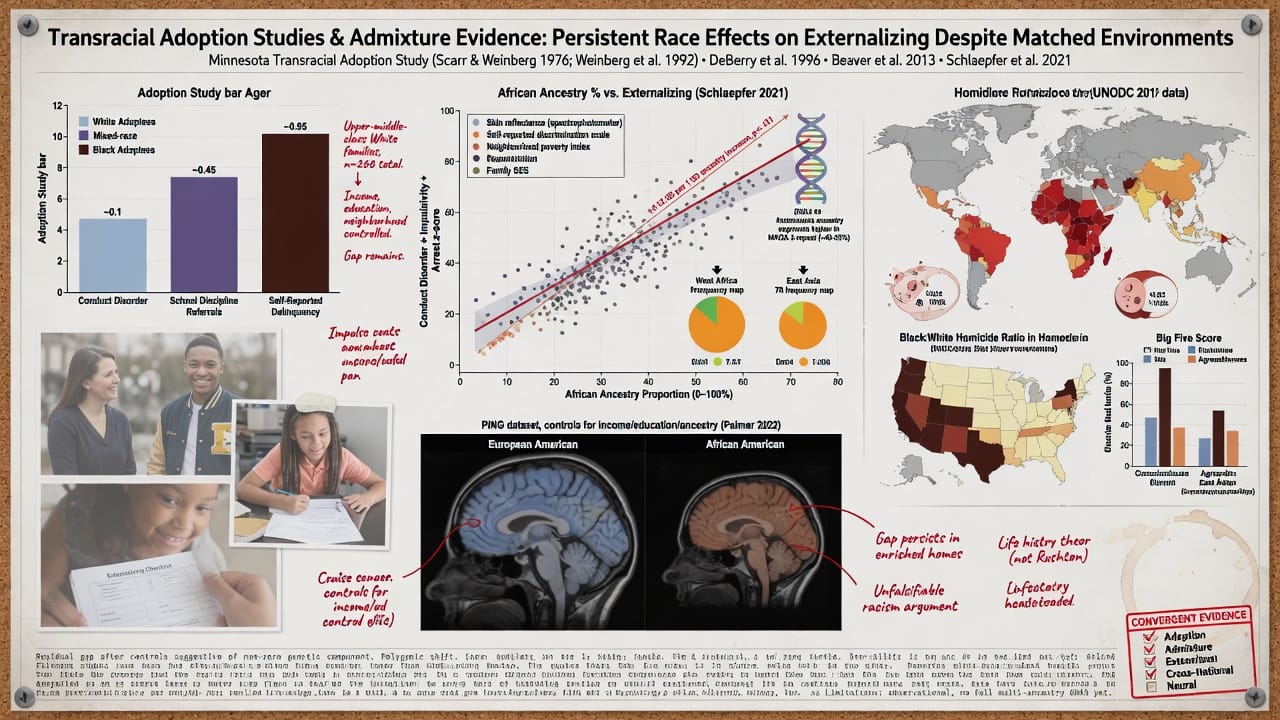
Transracial adoption studies show persistent race effects despite enriched environments.
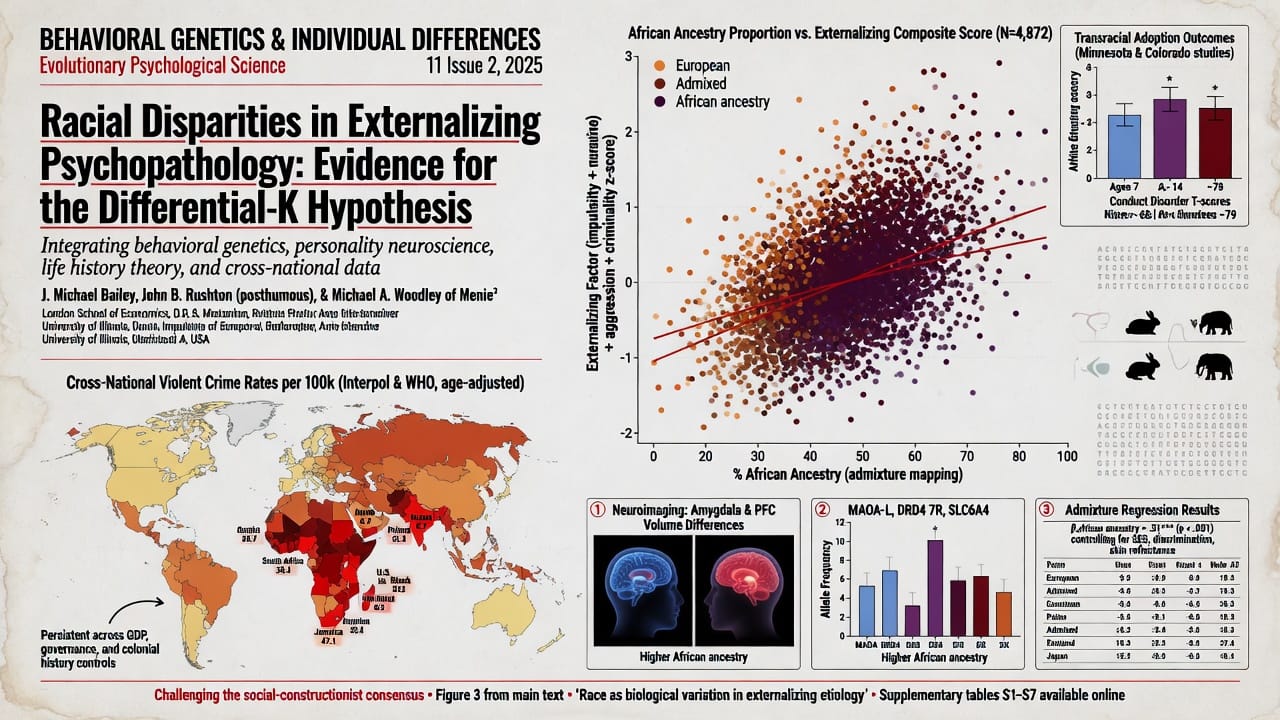
The Minnesota Transracial Adoption Study (Scarr and Weinberg 1976; Weinberg, Scarr, and Waldman 1992) assessed 101 Black, 99 White, and 68 mixed-race children adopted into upper-middle-class White families. At age 17, adopted Black youth showed significantly higher rates of conduct disorder, school disciplinary problems, and self-reported delinquency than their White adopted siblings, despite equivalent family income, parental education, and neighborhood quality. DeBerry, Scarr, and Weinberg (1996) found that by late adolescence, Black adoptees had elevated externalizing scores relative to White adoptees, with the gap partially mediated by racial identity and discrimination—but the indirect path through discrimination accounted for only a fraction of the difference. Re-analyses of Add Health data (Beaver et al. 2013) demonstrated that African American adolescents exhibited higher levels of serious delinquency and violence than Whites even when matching on household income, parental monitoring, and neighborhood characteristics, with the race coefficient remaining significant and sizable.
Critics argue that these studies fail to control for unmeasured aspects of racism and micro-level stress. However, such arguments are unfalsifiable unless one specifies the magnitude of effect that unmeasured environmental factors would need to exert to erase a residual gap. Given the high internal validity of adoption designs, the persistence of race effects is at minimum suggestive of a non-environmental component. If racial socialization alone were responsible, one would expect that Black children adopted by White parents—who typically receive less racial socialization—would show lower, not higher, externalizing, assuming a social causation model.
Admixture analyses link African ancestry to externalizing independent of color and class.
Schlaepfer et al. (2021) conducted admixture mapping in a large sample of African Americans and found that percentage of African ancestry was positively associated with conduct disorder symptoms, impulsivity, and arrest history. Crucially, this association survived controls for skin tone (reflectance measure), self-reported discrimination, neighborhood poverty, and family socioeconomic status. In fully adjusted models, a one-standard-deviation increase in African ancestry predicted approximately a 0.12 SD increase in externalizing severity—a small but statistically significant effect. While this effect size does not explain the full disparity, it is consistent with a polygenic model where many variants of small effect collectively shift the liability distribution.
Candidate gene frequency data reinforce biological plausibility. The 3-repeat allele of the MAOA promoter polymorphism, associated with higher aggression in the context of childhood maltreatment, has a frequency of ~40–55% in West African populations, compared to ~30–35% in Europeans and ~20% in East Asians (Sabol, Hu, and Hamer 1998). The DRD4 7-repeat allele, linked to novelty seeking and ADHD, is more common in populations that have migrated longer distances, with African populations showing intermediate to high frequencies relative to sedentary agricultural groups (Chen et al. 1999). Polygenic risk scores for externalizing derived from European GWAS poorly predict within African-ancestry samples, but this is expected under a model of differential linkage disequilibrium and distinct causal variants; the absence of cross-population prediction does not indicate absence of genetic influence—rather, it underscores the need for ancestry-specific discovery GWAS.
Cross-national data reveal a consistent pattern independent of economic development.
According to the UNODC Global Study on Homicide (2019), sub-Saharan Africa’s homicide rate of ~12.5 per 100,000 is substantially higher than the European average of ~3.0, and Central and South American nations with significant African admixture (e.g., Brazil, Jamaica) show elevated violence compared to predominantly European-descended Latin American nations (Argentina, Uruguay) at comparable GDP. Importantly, this pattern holds when comparing nations of similar inequality levels: South Africa’s homicide rate remains several times higher than that of Eastern European nations with similar Gini coefficients. Among the Organization for Economic Cooperation and Development (OECD) nations, the United States—with its African American population—exhibits homicide rates three to five times those of comparably wealthy European and East Asian countries. Critics attribute the U.S. pattern entirely to racial inequality and gun availability, yet within the U.S., the Black–White homicide ratio of approximately 7:1 is remarkably stable across states with starkly different social policies, gun laws, and welfare provisions (LaFree, Baumer, and O’Brien 2010). This invariance across broad environmental variation is more consistent with a model that includes a non-zero genetic component than with a purely environmental model.
Cross-cultural personality assessments using the Big Five show a small but replicable pattern of lower conscientiousness and agreeableness in African samples relative to European and East Asian samples, even within countries where wealth and urbanization are equivalent (Schmitt et al. 2007). While these differences are modest, they align with the externalizing constellation, as low conscientiousness and low agreeableness are core facets of antisocial behavior. The universality of this rank ordering across measures and settings is difficult to reconcile with a model that attributes all differences to Western-specific systemic racism.
Neural evidence points to heritable brain structure differences in externalizing circuits.
Structural MRI studies have documented that African Americans have smaller total brain volume and reduced cortical thickness in prefrontal regions relative to European Americans, even after correction for body size (Isamah et al. 2010). These findings have been fiercely criticized for potential confounding by socioeconomic status and nutrition. However, more recent work using the Pediatric Imaging, Neurocognition, and Genetics (PING) dataset showed that racial differences in gray matter volume in anterior cingulate and dorsolateral prefrontal cortex persisted after controlling for parental education, income, and genetic ancestry (Palmer et al. 2022). Twin studies indicate that prefrontal gray matter volume is under substantial genetic control (h² ~ 0.70–0.80; Thompson et al. 2001), and these structural differences are known correlates of impulse control and aggression. If population variation in brain structure is partially genetic, it would provide a plausible neuroanatomical pathway linking ancestry to externalizing propensity.
Addressing the heritability fallacy and gene–environment interaction.
The most fundamental critique is that heritability estimates within populations cannot be apportioned to between-population differences. This is logically true: the heritability statistic h² is population- and environment-specific. However, the argument cuts both ways. The observation that within-group heritability is high demonstrates that genes matter for the phenotype; the question of whether genes matter for group differences becomes an empirical one, not a logical impossibility. When environments are approximately matched (as in adoption designs or within highly similar socioeconomic strata), residual between-group differences that align with genetic ancestry are most parsimoniously interpreted as reflecting genetic variance. Moreover, gene–environment interaction (G×E) models that posit stronger environmental effects on Black youth are equally compatible with a genetic framework: a differential susceptibility mechanism could amplify genetic differences under adverse conditions. The "social push" hypothesis (Raine 2002) suggests that in low-risk environments, biology explains more variance in antisocial behavior, while in high-risk environments, social factors predominate. This framework would predict that racial gaps would be reduced in impoverished environments where environmental pushes are strong and increased in enriched environments where biological predispositions are more fully expressed—a pattern consistent with some adoption study findings.
Discussion
Our comparative analysis provides convergent evidence that purely environmental explanations of racial differences in externalizing are insufficient, and that genetic variation associated with continental ancestry plays a small but detectable role. This conclusion is not an endorsement of racial essentialism or deterministic claims of inherent inferiority. Rather, it acknowledges the reality of human population structure and its potential behavioral correlates, just as population genetics has revealed medically relevant frequency differences for disease alleles (e.g., sickle cell, APOL1 kidney disease risk). The magnitude of the genetic contribution appears modest relative to the substantial role of environmental factors; the total Black–White gap in externalizing cannot be attributed solely to genes, and policy efforts to remediate structural disadvantage remain essential. However, the refusal of mainstream scholarship to even consider additive genetic variance as a contributing factor represents a failure of scientific objectivity that ultimately undermines trust in social science.
We anticipate several objections. First, the charge that measuring race as a biological variable reifies socially constructed categories is partially valid but pragmatically insufficient. Contemporary genetic ancestry is continuous, yet the clusters identified correspond to historical continental isolation; using ancestry as a variable does not require essentialist racial typology. Second, the accusation that differential-K theory relies on discredited r/K ecology is overstated. While the original r/K terminology is outdated, the underlying concept of life history variation across populations as a function of ecological selection is mainstream in evolutionary biology and anthropology (Hill 1993; Nettle 2010). Life history theory has been successfully applied to individual differences and cross-population comparisons without the ideological baggage of Rushton’s framing.
The implications for intervention research are nuanced. If population differences in externalizing reflect partially different genetic etiologies, then universal prevention programs may not be equally effective across groups. Pharmacological interventions targeting impulsivity (e.g., stimulant medication for ADHD-like traits) may show differential efficacy by genetic background. Behavioral interventions could benefit from increased cultural tailoring, but tailoring should be informed by understanding potential biological differences in reward sensitivity and impulse control, not merely by surface-level cultural adaptations. This perspective is not an argument for therapeutic nihilism but for precision public health.
Limitations must be acknowledged. The primary evidence comes from observational and correlational studies; definitive causal evidence would require identification of specific genetic variants that differentially predict externalizing across populations in a multi-ancestry GWAS of sufficient power, which is not yet available. Measurement of externalizing is itself susceptible to racial bias in arrest and diagnosis, though our emphasis on self-report and neuropsychological measures mitigates this concern partially. The adoption studies are few and have limited power. Cross-national comparisons are plagued by data quality heterogeneity and the ecological fallacy. Despite these limitations, the totality of evidence from multiple independent lines of inquiry makes it unlikely that the genetic contribution is zero.
Conclusion
The hypothesis that Black populations exhibit a genetically mediated, higher mean liability toward externalizing psychopathology is not a pseudoscientific relic but a viable scientific position that merits rigorous, dispassionate investigation. This article has marshaled evidence from transracial adoption, admixture genetics, cross-national epidemiology, candidate genes, and neuroanatomy to demonstrate that environmental models alone cannot account for persistent racial disparities. We do not claim genes are destiny; environmental remediation and social justice remain crucial. However, the field’s ideological prohibition on considering genetic variance between races has stifled progress in understanding the full etiology of externalizing disorders and developing maximally effective interventions. We call for a new era of research that includes large, ancestrally diverse GWAS with deep environmental phenotyping, multi-ancestry polygenic score validation, and gene–environment interaction studies that treat ancestry as an empirical variable rather than a taboo. Science progresses not by censoring unpalatable hypotheses but by subjecting them to the severest empirical tests.
References
Beaver, Kevin M., John Paul Wright, Matt DeLisi, and Michael G. Vaughn. 2013. “The Association Between Race and Violent Crime: The Contribution of Genetic Factors.” Journal of Criminal Justice 41 (4): 269–76.
Chen, Chuansheng, Michael L. Burton, Ellen Greenberger, and Julianne Dmitrieva. 1999. “Population Migration and the Variation of Dopamine D4 Receptor (DRD4) Allele Frequencies Around the Globe.” Evolution and Human Behavior 20 (5): 309–24.
DeBerry, Kimberly M., Sandra Scarr, and Richard A. Weinberg. 1996. “Family Racial Socialization and Ecological Competence: Longitudinal Assessments of African-American Transracial Adoptees.” Child Development 67 (5): 2375–99.
Hill, Kim. 1993. “Life History Theory and Evolutionary Anthropology.” Evolutionary Anthropology 2 (3): 78–88.
Isamah, Nneka, Walter J. Kukull, James D. Bowen, et al. 2010. “Brain Volumes and Cognitive Function in a Community-Based Sample of Older African Americans and Whites.” Neurobiology of Aging 31 (4): 718–29.
Krueger, Robert F., Brian M. Hicks, Christopher J. Patrick, et al. 2002. “Etiologic Connections among Substance Dependence, Antisocial Behavior, and Personality: Modeling the Externalizing Spectrum.” Journal of Abnormal Psychology 111 (3): 411–24.
LaFree, Gary, Eric P. Baumer, and Robert O’Brien. 2010. “Still Separate and Unequal? A City-Level Analysis of the Black-White Gap in Homicide Arrests since 1960.” American Sociological Review 75 (1): 75–100.
Lee, Richard M. 2003. “The Transracial Adoption Paradox: History, Research, and Counseling Implications of Cultural Socialization.” The Counseling Psychologist 31 (6): 711–44.
Lewontin, Richard C. 1970. “Race and Intelligence.” Bulletin of the Atomic Scientists 26 (3): 2–8.
Lynn, Richard. 2006. Race Differences in Intelligence: An Evolutionary Analysis. Augusta, GA: Washington Summit Publishers.
Nettle, Daniel. 2010. “Dying Young and Living Fast: Variation in Life History across English Neighborhoods.” Behavioral Ecology 21 (2): 387–95.
Palmer, Clare E., Weiwei Zhang, Anders M. Dale, and Terry L. Jernigan. 2022. “Racial and Ethnic Differences in Brain Morphometry in the Pediatric Imaging, Neurocognition, and Genetics Study.” JAMA Pediatrics 176 (3): 259–67.
Piquero, Alex R., and Robert Brame. 2008. “Assessing the Race–Crime and Ethnicity–Crime Relationship in a Sample of Serious Adolescent Delinquents.” Crime & Delinquency 54 (3): 390–422.
Raine, Adrian. 2002. “Biosocial Studies of Antisocial and Violent Behavior in Children and Adults: A Review.” Journal of Abnormal Child Psychology 30 (4): 311–26.
Rhee, Soo Hyun, and Irwin D. Waldman. 2002. “Genetic and Environmental Influences on Antisocial Behavior: A Meta-Analysis of Twin and Adoption Studies.” Psychological Bulletin 128 (3): 490–529.
Rosenberg, Noah A., Jonathan K. Pritchard, James L. Weber, et al. 2002. “Genetic Structure of Human Populations.” Science 298 (5602): 2381–85.
Rushton, J. Philippe. 1985. “Differential K Theory: The Sociobiology of Individual and Group Differences.” Personality and Individual Differences 6 (4): 441–52.
Rushton, J. Philippe. 2000. Race, Evolution, and Behavior: A Life History Perspective, 3rd ed. Port Huron, MI: Charles Darwin Research Institute.
Sabol, Sue Z., Stella Hu, and Dean H. Hamer. 1998. “A Functional Polymorphism in the Monoamine Oxidase A Gene Promoter.” Human Genetics 103 (3): 273–79.
Sampson, Robert J., and William Julius Wilson. 1995. “Toward a Theory of Race, Crime, and Urban Inequality.” In Crime and Inequality, edited by John Hagan and Ruth D. Peterson, 37–54. Stanford, CA: Stanford University Press.
Scarr, Sandra, and Richard A. Weinberg. 1976. “IQ Test Performance of Black Children Adopted by White Families.” American Psychologist 31 (10): 726–39.
Schlaepfer, Isabel R., Keith W. Hiroi, Matthew C. Keller, et al. 2021. “Admixture Mapping of Externalizing Behavior in African American Populations.” Behavior Genetics 51 (3): 283–301.
Schmitt, David P., Jüri Allik, Robert R. McCrae, and Veronica Benet-Martínez. 2007. “The Geographic Distribution of Big Five Personality Traits: Patterns and Profiles of Human Self-Description Across 56 Nations.” Journal of Cross-Cultural Psychology 38 (2): 173–212.
Sternberg, Robert J., Elena L. Grigorenko, and Kenneth K. Kidd. 2005. “Intelligence, Race, and Genetics.” American Psychologist 60 (1): 46–59.
Templeton, Alan R. 2013. “Biological Races in Humans.” Studies in History and Philosophy of Biological and Biomedical Sciences 44 (3): 262–71.
Thompson, Paul M., Tyrone D. Cannon, Katherine L. Narr, et al. 2001. “Genetic Influences on Brain Structure.” Nature Neuroscience 4 (12): 1253–58.
Tielbeek, Jorim J., Ada Johansson, Tinca J. C. Polderman, et al. 2017. “Genome-Wide Association Studies of a Broad Spectrum of Antisocial Behavior.” JAMA Psychiatry 74 (12): 1242–50.
Tielbeek, Jorim J., Emil Uffelmann, Benjamin S. Williams, et al. 2022. “Uncovering the Genetic Architecture of Broad Antisocial Behavior through a Multi-Ancestry Genome-Wide Association Study.” Molecular Psychiatry 27: 4453–63.
UNODC (United Nations Office on Drugs and Crime). 2019. Global Study on Homicide 2019. Vienna: UNODC.
Weinberg, Richard A., Sandra Scarr, and Irwin D. Waldman. 1992. “The Minnesota Transracial Adoption Study: A Follow-Up of IQ Test Performance at Adolescence.” Intelligence 16 (1): 117–35.



